How to make male sperm
To understand how one might make female sperm, one must first understand how to make male sperm (and then how to change male sperm). Also, knowing how males makes sperm reveals interesting insights into the genetic relationship between men and women (for example, one might SRYly argue, by considering the few genes on the Y chromosome, that men are just women with sperm factories). Making sperm can be viewed as a six step process - the first four steps that mostly occur in the male embryo that prepare generic diploid germ cells, and the last two steps that mostly occur in the post-puberty male that convert germ cells into haploid sperm (think of germ cells as pre-sperm cells):
- CREATE - germ cells are created in the embryo yolk sac
- LOCATE - germ cells migrate to the proto-testicals
- COOK - germ cells expand in number
- SIMMER - germ cells stop expanding and wait for puberty
- DAZZINGLY SPLIT - post puberty, diploid germ cells are converted to haploid pre-sperm
- ASSEMBLE - the pre-sperm have tails added to become sperm
- DELIVER - the sperm find their way to the egg
- CLEANING - cleaning out bad sperm
- KLINEFELTER - sperm from non-mosaic 47,XXY males
The biology Web site www.maxamin.com has some nice animations on genetics, including two of interest to sperm production: the meiotic process of Spermatogenesis, and Stages of Meiosis.
Professor Danton O'Day of the University of Toronto has a nicely illustrated overview of the basics of sperm production. Click here.
The American Society of Andrology has an excellent online handbook to the male reproductive system and sperm.
| TIMING | MALE GERM CELLS |
|---|---|
| 3 weeks post-fertilization | germ cells on wall of yolk sac |
| 4 weeks post-fertilization | germ cells start migrating |
| 5 weeks post-fertilization | germ cells arrive at primitive gonads |
| 6 weeks post-fertilization | germ cells in genital ridges |
| 7 weeks post-fertilization | male sex organs start forming |
| 12 weeks post-fertilization | sperm are imprinted ??? |
| 20 weeks post-fertilization | testes develop |
CREATE - germ cells are created in the embryo yolk sac
There are certain cells in the body used to make other, more specialized cells. These cells are called "stem cells". For example, blood stem cells are a generalized cell that the body converts into red blood cells, white blood cells and other cells appearing in the blood. One type of stem cell is the "germ cell", which is used to make sperm and eggs. For males, "germ cells" are a kind of pre-sperm. The National Institutes of Health has a nice introduction to stem cells.
Germ cells first appear in the embryo about three weeks after fertilization. At this stage, the embryo is about the size of a grain of rice???, has no sex, and much of the embryo is a yolk sac where the germ cells are initially found.
About this point in time, inherited imprinting (imprinting is discussed below) is erased.
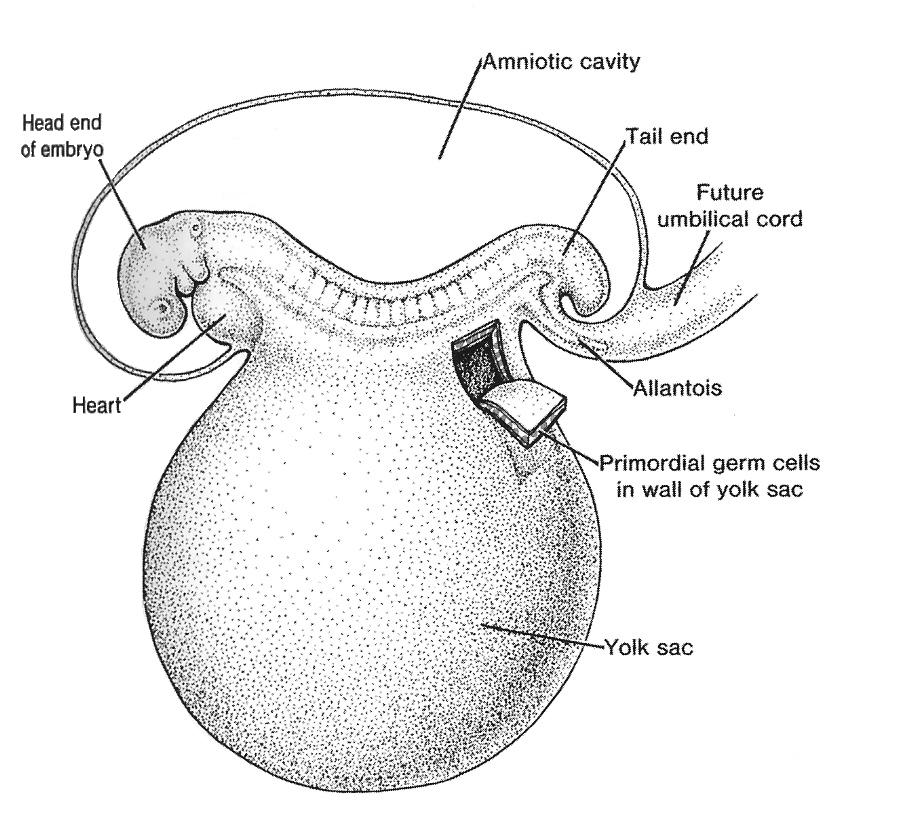
Picture credit: Langman's Medical Embryology [1995], page 11]
The University of Calgary's Virtual Embryo project has a nice introduction to germ cells.
LOCATE - germ cells migrate to the proto-testicals
About one week after the germ cells appear in the yolk sac, they start traveling out of the yolk sac (the yolk sac eventually disappears) and migrate to the genital ridge area of the embryo (see Figure (B) above), reaching the genital ridge around the sixth week after fertilization. Here the germ cells trigger the development of the sex organs around the seventh week after fertilization - the embryo is structurally sexless until this point.
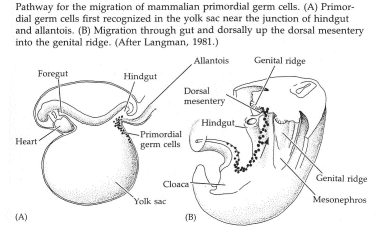
COOK - germ cells expand in number
The germ cells follow along in the creation of the testicles, ending up in the region of the testicles known as the seminiferous tubules, a spaghetti-like structure where sperm are manufactured. The germ cells continue to expand in number.
At the earliest stages of embryo development, the first five to seven weeks or so, the gonads of individuals do not exhibit morphological differences (Figures 2A and 2B from the app) and are essentially female, for example, with a double set of sex ducts. In the male gonad, primordial germ cells in the cortex begin to degenerate while the testis cords in the center of the gonad continue to develop into the seminiferous tubules. The gonad is converted into a testis. In the female gonad, the central testis cords disappear, while the oocytes in the peripheral cortex survive and become surrounded by follicle cells; the gonad is developing into an ovary.
Mammalian embryos are initially "sexless", with the default pathway being ovarian differentiation (glibly, men from ovaries, not women from ribs). The presence of the SRY/TDF gene (sex-determining region Y / testis-determining factor gene) on the Y chromosome (for primates, for example) forces the gonad to become male. The DSS (dosage-sensitive sex reversal) locus also suggests being female as the default case for mammals, with SRY proposed to override one or more genes in DSS, and with two copies of DSS overwhelming SRY, causing male-to-female sex reversal. For males, epithelial somatic cells of the bipotent gonadal primordium differentiate into Sertoli cells, and this in turn triggers testis development.
SIMMER - germ cells stop expanding and wait for puberty
At some point during the lifetime of the embryo, the germ cells stop expanding in number, and go into a form of hibernation, waiting for the male to reach puberty.
Through the embryo and fetal stages, and up until puberty, the male's germ cells are diploid. Male germ cells do not split their DNA until after puberty - that is, adult males still have diploid germ cells. This is an important distinction between men and women - in the embryo, female diploid germ cells divide into haploid cells - so that adult females have no diploid germ cells (this is addressed on the making female sperm Web page). Click here for an introduction to and pictures of human chromosomes
DAZZINGLY SPLIT - post puberty, diploid germ cells are converted to haploid pre-sperm
SPERMATOGENESIS
Once a male reaches puberty, he starts making sperm. The
germ cells, which had been in a rest state since the male's birth, start
being acted upon to produce sperm. The first stage of this production is
known as spermatogenesis, where germ cells are used to produce
spermatids (basically sperm without tails). The second stage,
spermiogenesis, is where the spermatids are transformed into
full sperm with tails. Spermiogenesis is discussed below.
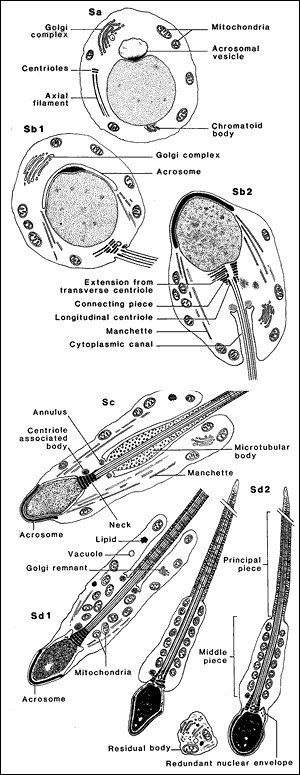
Spermatogenesis has two acts. In the first act, the germ cells cast off daughter cells known as spermatogonia. The spermatogonia grow in number through mitosis (cell devision where the daughter cells have the same number of chromosomes as the original cell). Then the spermatogonia split in half through meiosis, forming spermatocytes and then spermatid. The spermatid are haploid, having half of the chromosomes of the original germ cells. Click here for an introduction to mitosis and meiosis.
Click here for two excellent animations of Mitosis and Meiosis, part of John Kyrk's excellent Web site on cell biology animation.
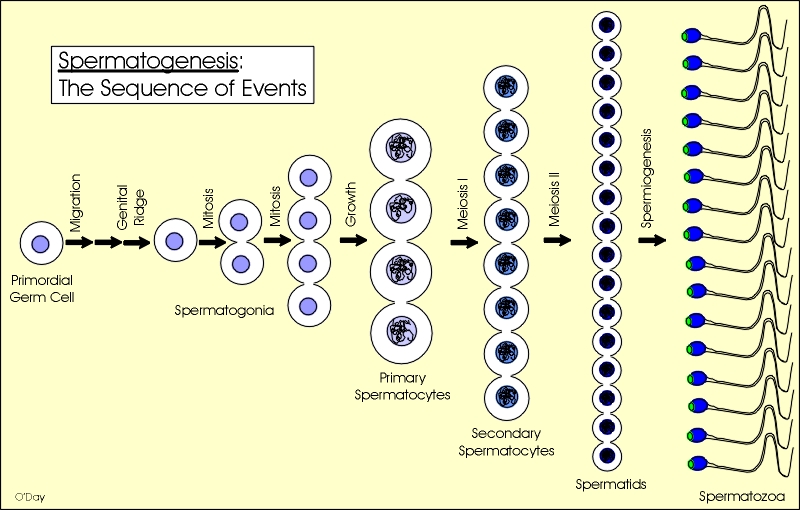
Picture credit:
Prod. Danton O'Day, University of Toronto.
{kind=link}
Two very important transformations occurs during this step. First, some of the genes in the chromosomes are imprinted to indicate that the genes came from a male. Similarly, when a woman's eggs are being created, some of the genes in the egg's chromosomes are imprinted to indicate that the genes came from a female (and the imprinting is different from that of male imprinting). When the genes in a sperm combine with the genes in an egg, they have to be imprinted differently, otherwise the embryo either aborts at some point or has a damaged development. Imprinting will be discussed further in this Web site.
Imprinting has been recognized, if not understood, for a very long time. About 3000 years ago, people breeding mules observed that if you breed a donkey with a mare, the resulting offspring are mules. However, if you breed a donkey with a stallion, the offspring are hinnies. Hinnies differ from mules by having stronger legs, shorter ears and a thicker tail and mane. It was apparent even then that males and females contributed to their children slightly differently.
Second, during meiosis, the X and Y chromosomes separate a bit from the other chromosomes and form an XY body. XY bodies will be discussed further in the section on X-inactivation.
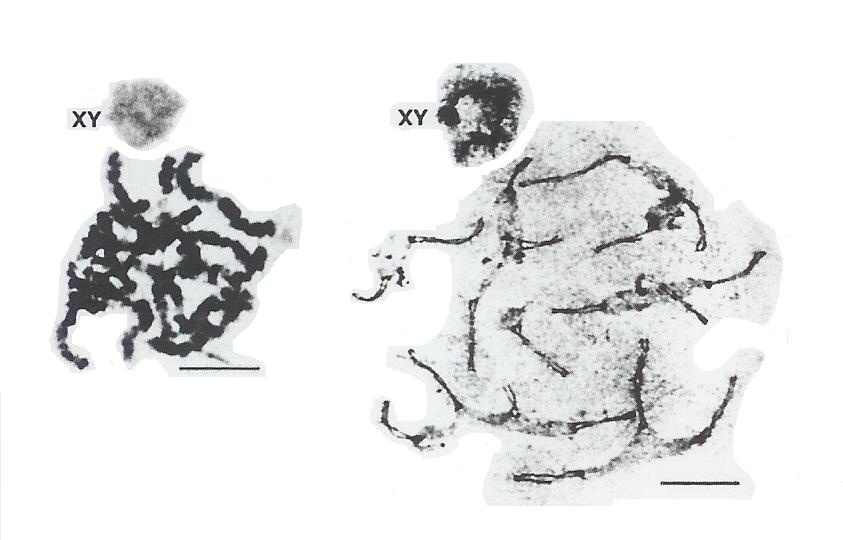
Picture credit: Nature Genetics [Nov 1992], page 192
The University of Calgary's Virtual Embryo project has a nice introduction to spermatogenesis.
ASSEMBLE - the pre-sperm have tails added to become sperm
SPERMIOGENESIS
The end result of the DAZZINGLY SPLIT stage is the formation of round
spermatid, i.e. sperm cells with no tails. Interestingly, experiments
have shown that round and elongated spermatids can be injected into eggs
to achieve fertilization, which helps men who are unable to completely
form sperm. Round spermatid can also be cultured in vitro to mature
their development, to make them more viable for IVF. That spermatid can
be used for fertilization implies that sufficient imprinting occurs in
the seminiferous tubule region, and that all changes to the spermatid
post-meiosis are mostly for insuring delivery of the sperm to the egg.
However, the safest route is to follow Mother Nature and form the tails. This process of converting spermatids to sperm is known as "spermiogenesis". The University of Calgary's Virtual Embryo project has a nice introduction to spermiogenesis. In spermiogenesis, the head of the sperm is reshaped, a tail is grown, and the components of the spermatid are rearranged. Even with a tail, fully-formed sperm are not that mobile. Rather as they leave the testis and travel through the epididymis they mature (sperm plasma membrane and some internal components change).
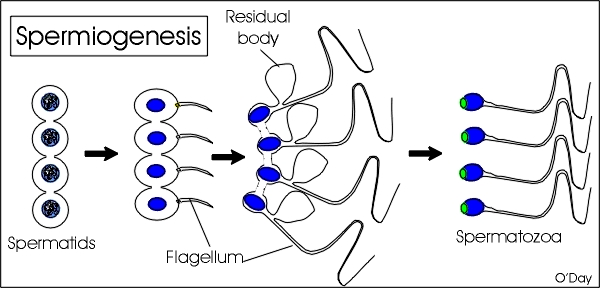
Picture credit:
Prod. Danton O'Day, University of Toronto.
{kind=link}
DELIVER - the sperm find their way to the egg
At this point, the sperm are almost ready for fertilizing eggs. The sperm leave the testicles, travelling through a tube - the vas deferens - which ends at the seminal vesicles. The seminal vesicles mix the sperm with a yellow fluid which is rich in fructose (a sugar) which give the sperm energy. From there, the sperm move to the prostate gland, which contributes some phosphate-related chemicals which help liquify the semen. At this point, the semen is ready to be ejaculated through the penis, with the sperm travelling through the penis' urethra, typically during sexual intercourse between a man and a woman (I don't know what kind of intercourse they call it when a man ejaculates into a cup at a fertility clinic).
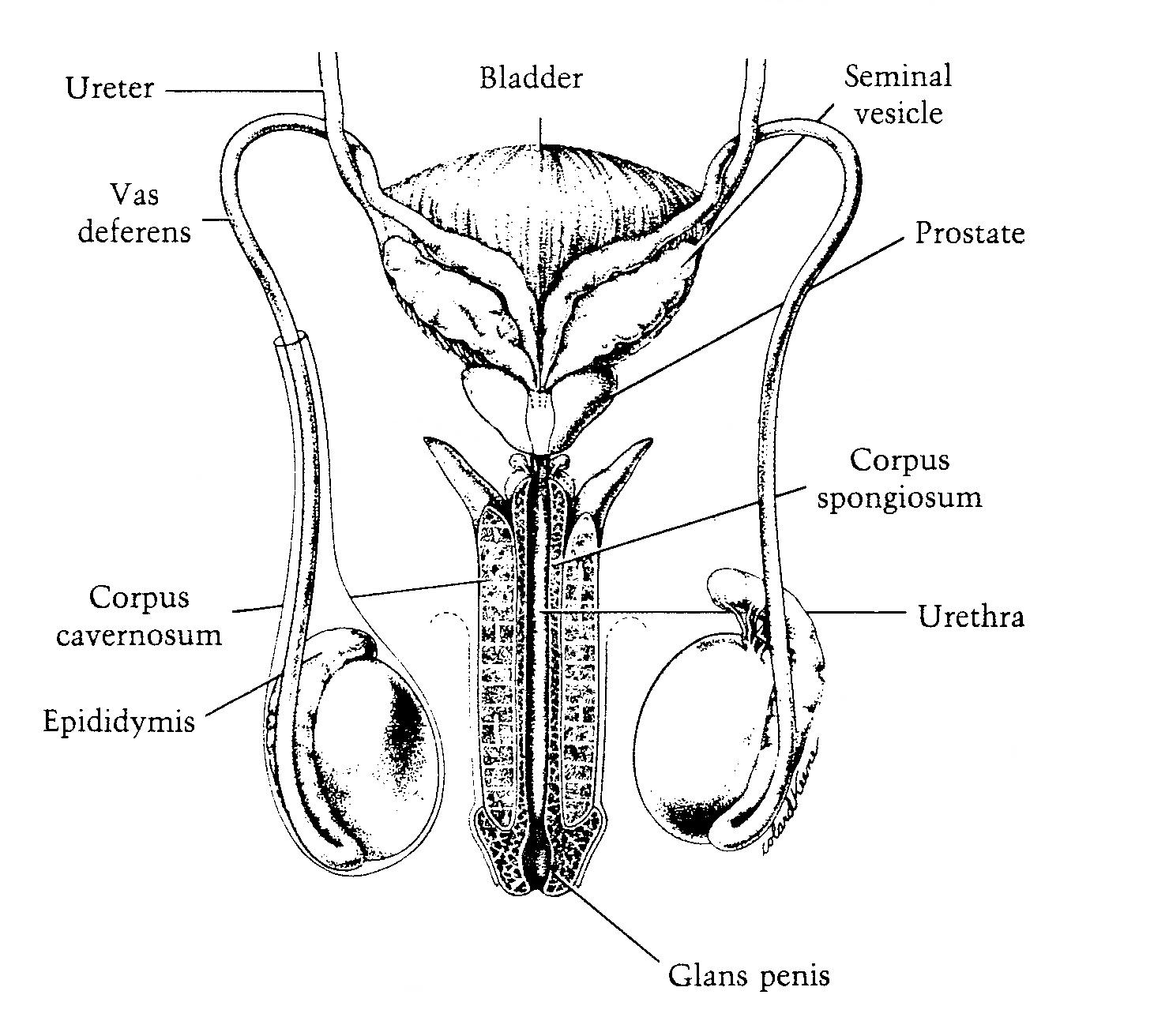
Picture credit: Atlas of Sperm Morphology (1989)
Religious authorities, in order to denigrate sexual pleasure somewhat, have associated the concepted of "uncleaness" with a man's ejaculation. While a bit messy (use a hot towel), a man's semen is, if anything, fairly nutritious (though I prefer donuts). Here is a table of chemicals found in semen.
TABLE OF SEMINAL PLASMAAfter ejaculation, as the sperm move through the woman's reproductive tract, they undergo capacitation where secretions alter the sperm plasma membrane that allows the sperm to attached to the egg.
Sperm face daunting odds for one of them to fertilize an egg. There are about 350 million sperm released during one ejaculation into the vagina, with upwards of 25 percent of them being defective. About one million sperm are able to reach the uterus. Contractions of muscles in the uterus move the sperm towards the Fallopian tubes, but only about one thousand sperm make it this far. Of these thousand sperm, only one will be lucky enough to be able to fuse with the egg.
In some cases where the man has a low sperm count, or sperm that are not as mobile as they should be, a sperm is extracted from the man's testicles, and directly injected into an egg (or, as mentioned above, a spermatid is extracted from the testicles and injected into an egg). This medical procedure is known as ICSI. This eliminates the need for the sperm to be capacitated.
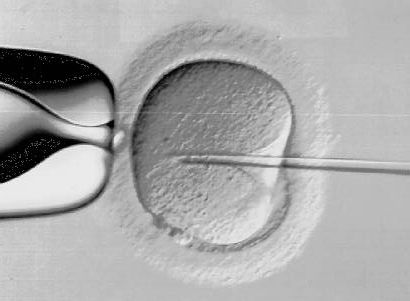
A nice introduction to this form of sperm injection, with some nice photographs of the sperm and egg at various, can be seen at: ICSI photographs.
CLEANING - cleaning out bad sperm
Men produce billions of sperm every month. Not surprisingly, some of the sperm are defective. To insure that defective sperm are not used to fertilize eggs, a whole industry has arisen that takes a sample of sperm and filters out defective sperm. Sperm can be defective in a variety of ways - have missing components, have two heads, have two tails, be mishaped, etc.
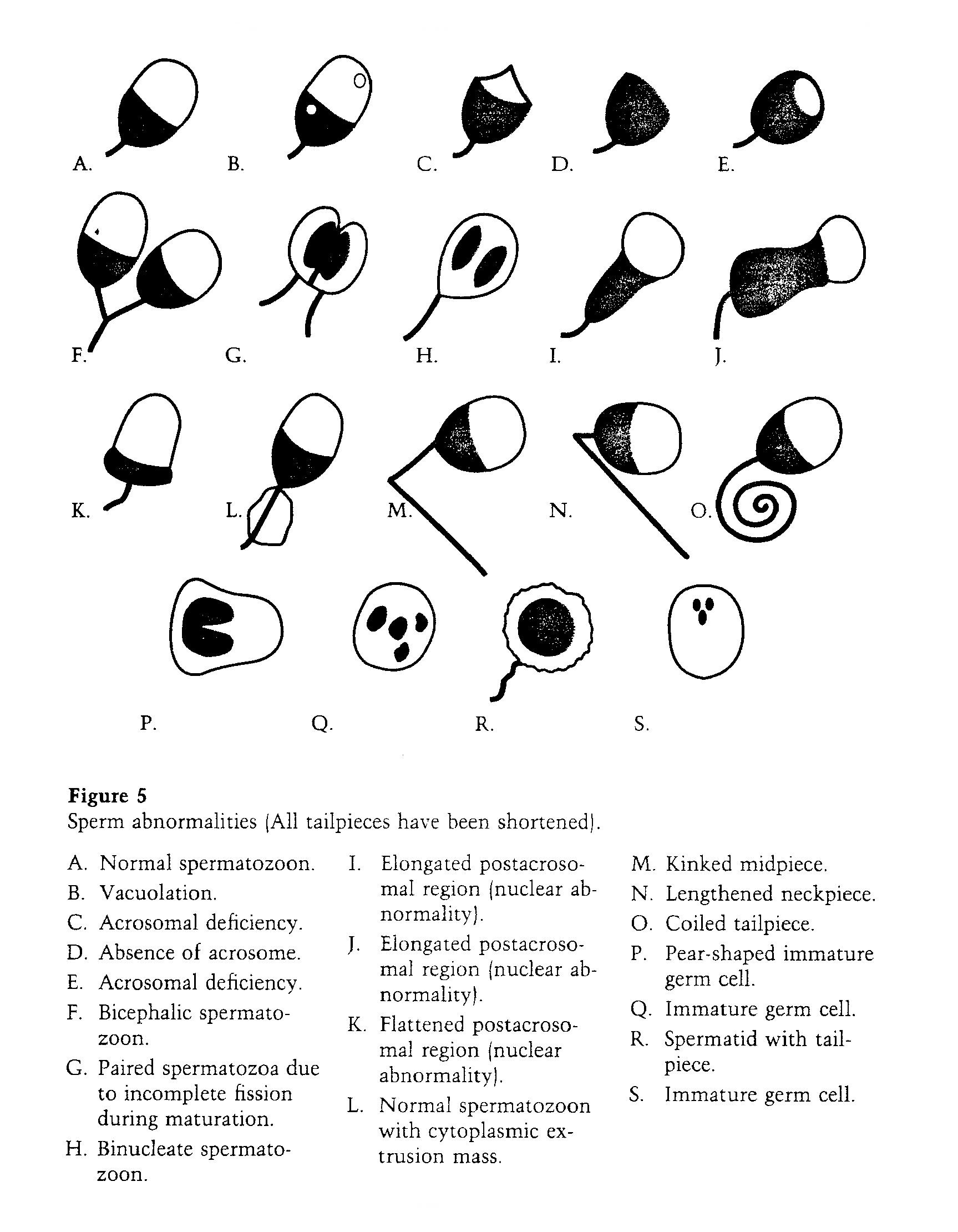
Defective Sperm - Picture credit:
Atlas of Sperm Morphology by Marilyn Adelman and Eileen Cahill, 1989
KLINEFELTER - sperm from non-mosaic 47,XXY males
There is a condition a small percentage of men are born with, the understanding of which will be very important to the key determining factor in achieving female sperm, i.e, meiosis of altered female diploid germ cells (females germ cells with 2 X and 1 Y chromosome). This conditions is known as Klinefelter's syndrome. What follows is an introduction to this condition and the current research to determine how such men are able to produce sperm (a process still not fully understood).
The vast majority of males can be genetically labeled as 46,XY males. That is, males have 46 chromosomes, 44 of which are mostly equivalent pairs, plus 2 sex chromosomes, 1 X and 1 Y. Females are genetically labelled as 46,XX females. In about one out of every 1000 male births, the male has an extra X chromosome, and is labelled either 47,XXY or 46,XY/47,XXY males. 47,XXY are labelled as "non-mosaic", meaning all of the cells in their body have 47 chromosomes. 46,XY/47,XXY males are labelled as "mosaic", meaning their body are a mixture of cells with 46 chromosomes and cells with 47 chromosomes.
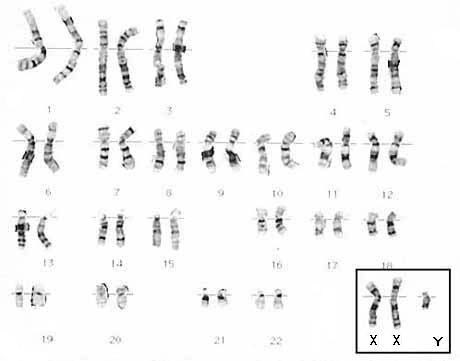
Picture credit:
Genetic Science Learning
Center, Department of Human Genetics, University of Utah.
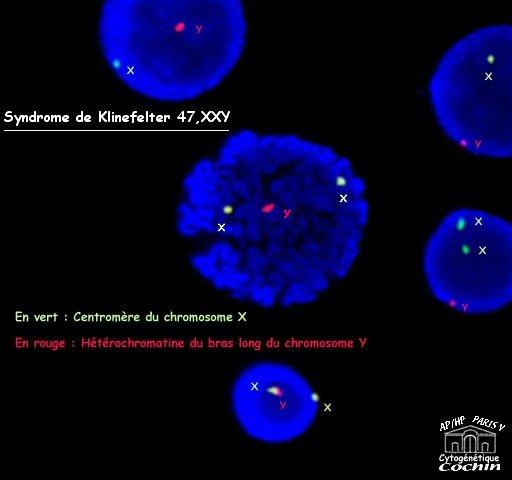
Picture credit:
Paris Virtual University
Males with Klinefelter Syndrome exhibit certain traits: they tend to be taller, have less body hair, and low sperm counts. Click here for introductions to Klinefelter's Syndrome and Understanding Klinefelter Syndrome.
It is not, and was not, surprising to learn that mosaic 46,XY/47,XXY males are able to make sperm, since there is likely to be enough sex organ cells and sperm cells with the normal gene count (46,XY) for the production of sperm to occur. The lower sperm counts can be attributed to the presence of the potentially disruptive 47,XXY cells. But what if the male is completely non-mosaic, with no 46,XY cells anywhere in the body, including no 46,XY sperm cells and no 46,XY sex organ cells. Do such non-mosaic males exist and can they make sperm (especially via meiosis)? This is a very important question to female sperm production since one way of making female could be to create 47,XXY female germ cells.
First, as far back as 1963, it was suspected that 47,XXY males could have viable sperm for fertilization (suggesting but not suggested that an XXY female germ cell could become sperm). The unresolved question is whether these non-mosaics are truly non-mosaics, and if not, whether 47,XXY germ cells would be meiotic in a purely 46,XY testicular environment (transplanted via Brinster), an environment with 46,XY Sertoli cells - the only somatic cells in direct contact with differentiating germ cells (Sertoli cells provide both physical and nutritional support for spermatogenesis, which occurs in the intercellular spaces between Sertoli cells).
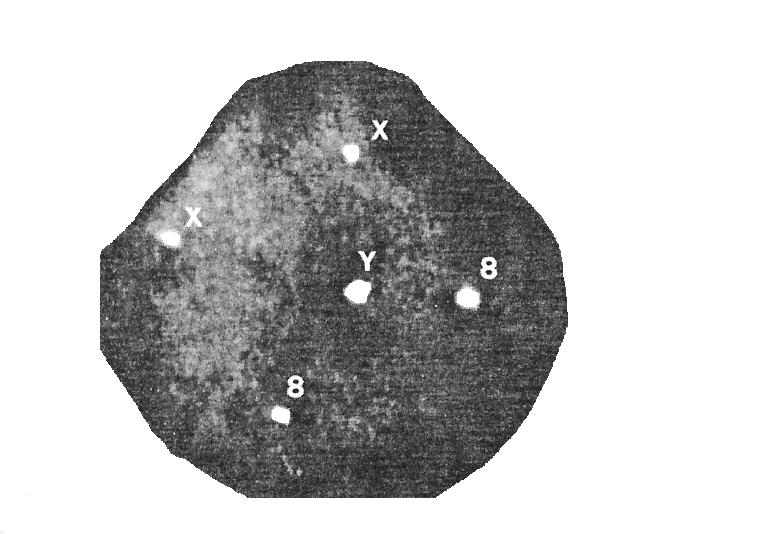
47,XXY Spermatogonia - Picture credit:
Carlo Foresta et.al.,J. Clin. Endo. Meta., v84 1999, 3807-10
Experiments in mice demonstrate in vitro MITOTIC preliferation of XXY germ cells (see the next section). This leaves the question if in vivo MEIOTIC proliferation can occur (for example in an XY testis) or in vitro meiotic proliferation, which addresses the problems of abnormal meiotic regulation in the XXY testis. This issue will be the main stumbling block to achieving female sperm.
Second, experiments with male mice bred to have a similar XXY condition have shown that such male mice again, while having low sperm counts, are able to produce sperm. Additionally, germ cells from such males are able to MITOTICALLY proliferate in vitro similar to germ cells from normal male mice.
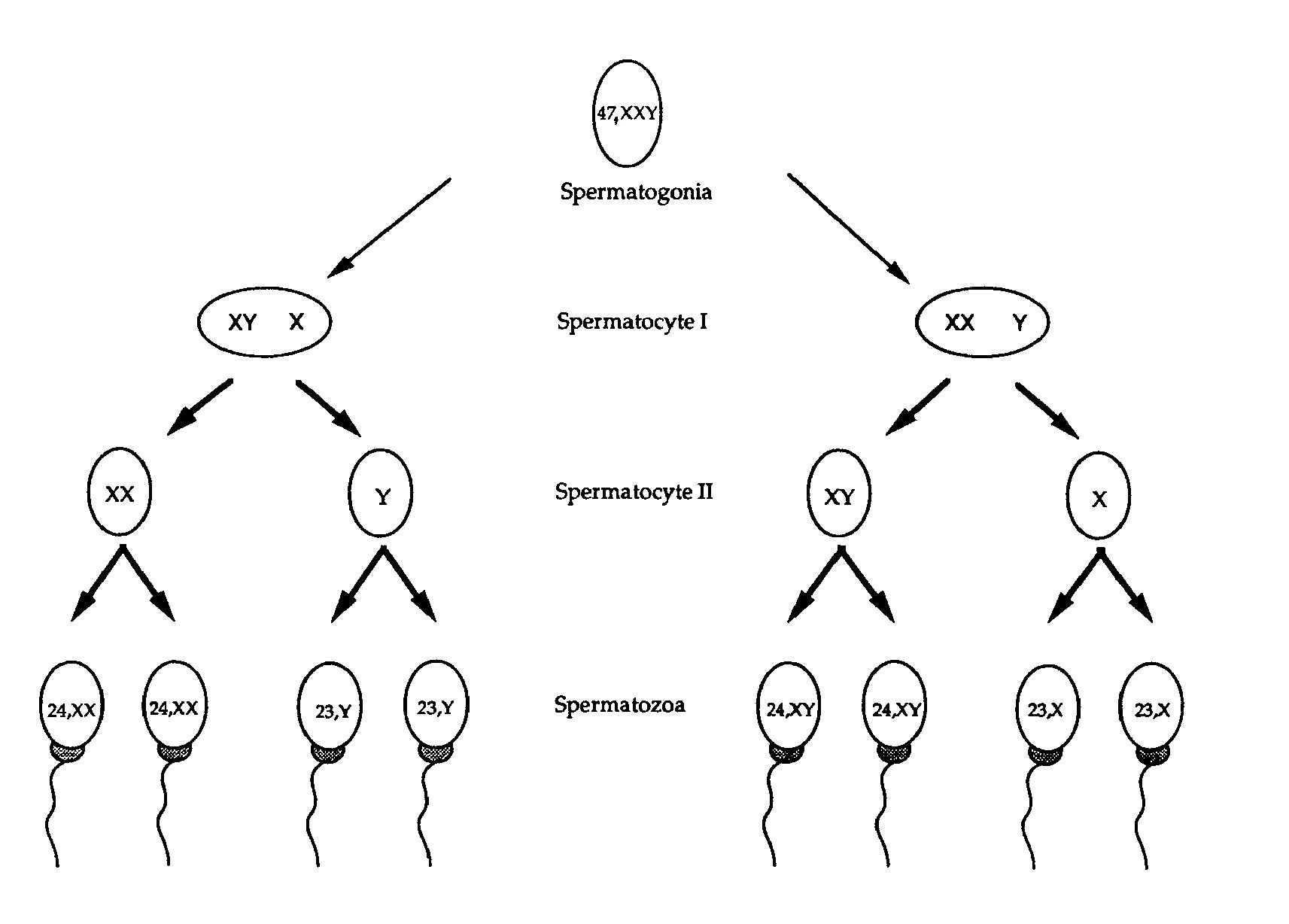
Possible XXY meiosis pathways
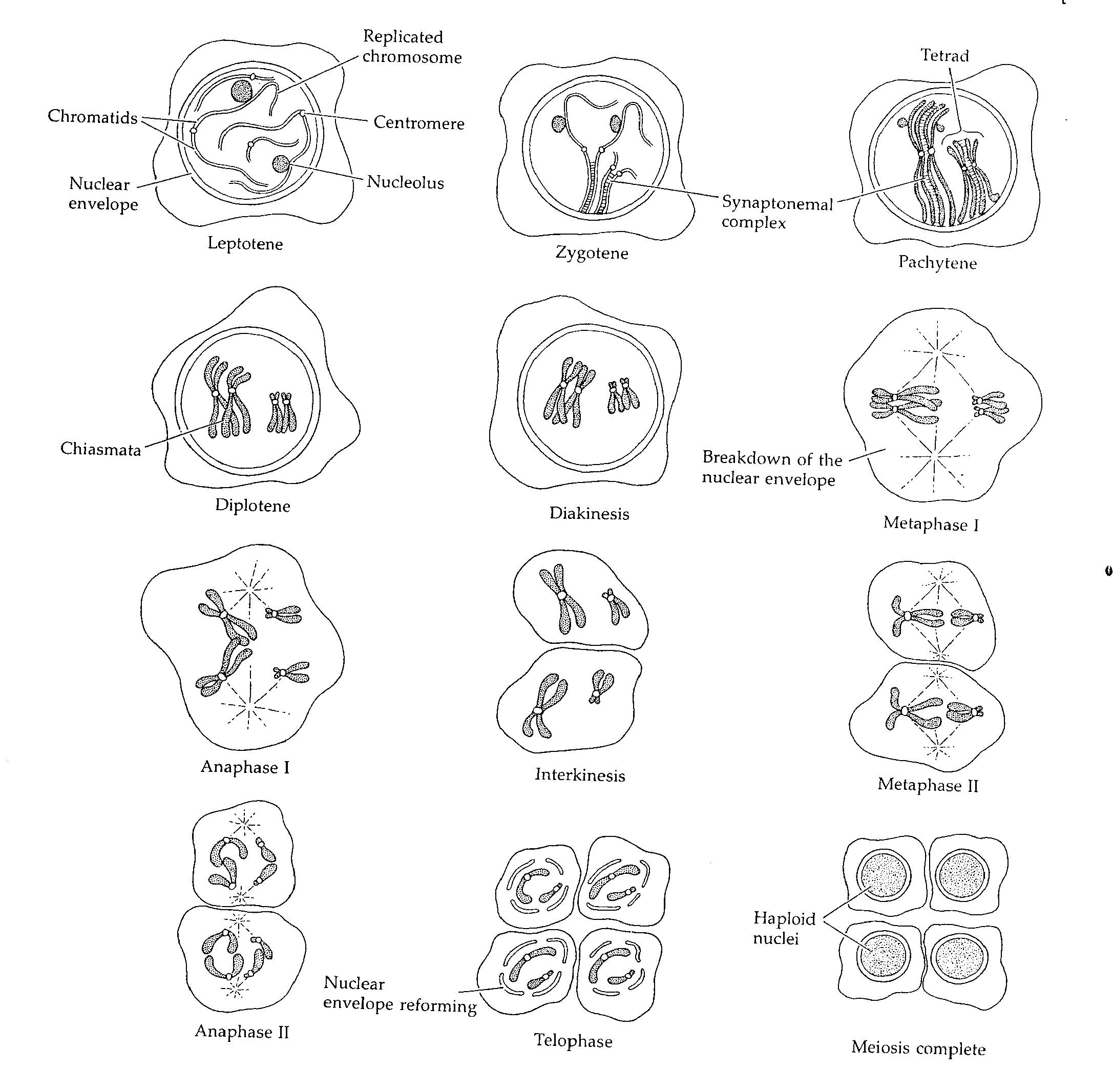
Stages of meiosis - Picture credit: Developmental Biology, Scott
Gilbert, 3rd edition, 793